Recently I have been thinking a lot about one of the most interesting topics: the origin of life. Specifically, the state of research on abiogenesis, the idea that life arose through purely natural processes without any intelligent guidance or intervention. I have also been thinking about the various viewpoints I have encountered over the years regarding abiogenesis.
The various viewpoints about abiogenesis can be categorized along something of a spectrum. Although the spectrum is not necessarily symmetrical and there are probably numerous different views, I found it helpful to break them down into seven different categories in a 3-1-3 distribution, with stronger support for abiogenesis at one end of the spectrum and stronger skepticism toward abiogenesis at the other end of the spectrum. These are of course approximations and simplifications, but I believe they provide a helpful way to assess where someone is coming from in their views on the topic.
I would ask readers to let us know (i) which viewpoint most closely approximates your position, and (ii) what additional clarifications or caveats, if any, you would add to more completely reflect your position. I also offer my take below each described view.
With that in mind, here are the categories.
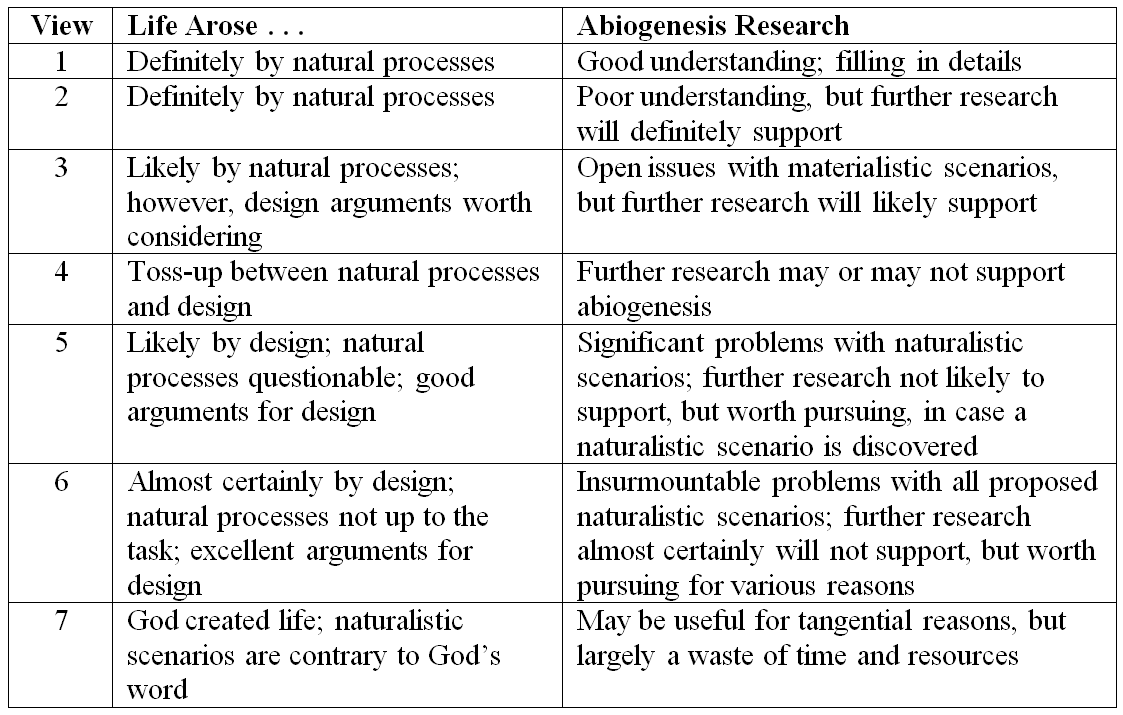
1. “Life arose through purely natural processes without any intelligent guidance or intervention. Furthermore, we have a pretty good understanding from a scientific standpoint of how this occurred, and at this point we are largely just filling in the details.”
Assessment. This viewpoint consists of: (i) a faith-based assertion of materialism, coupled with (ii) a misrepresentation of the scientific evidence. As a result, this is something of a minority position (given its looseness with the scientific facts). Yet it is a position we occasionally encounter, particularly when the misrepresentation serves some other propaganda need, such as Dawkins’ claim last year to a packed house in an NPR lecture series.
My Take. This is an untenable position that demonstrates a lack of intellectual and scientific integrity, and I have little patience for anyone putting forth such a deceptive claim.
2. “Life arose through purely natural processes without any intelligent guidance or intervention. Although we do not yet have a good idea from a scientific standpoint of how this occurred, in time we will uncover how it happened.”
Assessment. This viewpoint consists of: (i) a faith-based assertion of materialism, coupled with (ii) an acknowledgement of the lack of empirical evidence, but resting on hoped-for future discoveries that will confirm the materialistic position. This is a much more common position. Indeed, based on my years of experience following this issue, I would say that this is one of the most common positions, and is the position taken by most materialists as well as most scientific bodies or committees that have spoken on the issue.
My Take. This position is significantly better than the first, in that it acknowledges the current lack of viable naturalistic explanations. However, in practice it operates similarly to the first position, in that it avoids any discussion of alternate viewpoints and asserts that materialism is correct – we just need to wait for the evidence to catch up with the conclusion. As a result, it presents a veneer of intellectual integrity, but masks an underlying lack of objectivity.
3. “Life likely arose through purely natural processes without any intelligent guidance or intervention. We do not have a coherent story of how this could have occurred, and various open questions remain. Furthermore, there are some interesting arguments against a purely naturalistic scenario and in favor of intelligent guidance or design that should be considered. However, ultimately we will probably discover a plausible naturalistic scenario.”
Assessment. This is a more nuanced position that recognizes some of the weaknesses in the materialistic story and the existence of legitimate counter arguments. This position is also very common and would include individuals who are not necessarily design advocates, but who are either sympathetic to some of the design arguments or who are keenly aware of some of the limitations of purely naturalistic scenarios.
My Take. This position is more coherent and is, in my estimation, the farthest anyone could legitimately go in support of abiogenesis if they have taken an objective look at the evidence. It still relies on unspecified, hoped-for future discoveries, but allows for rational debate and discussion.
4. “Life may have arisen through purely natural processes or it might have arisen through intelligent guidance and intervention. We don’t know.”
Assessment. This is not really a position so much as a statement of a lack of position. This essentially functions as a temporary position for those who have not yet looked into the issue.
My Take. This position is rare and only temporary, but I include it for completeness and to show the centerline of the spectrum. Once people take time to look into abiogenesis, their viewpoint will inevitably shift in the direction of either #3 or #5 (in my experience, more often in the direction of #5).
5. “Life likely arose through intelligent guidance or intervention and not solely through natural processes. There are problems with essentially all naturalistic scenarios proposed to date. Furthermore, there are good reasons to think that a purely naturalistic scenario is infeasible, as well as strong arguments in favor of intelligent guidance or intervention. However, it remains possible that life arose through purely natural processes, and we should continue to search for plausible naturalistic scenarios.”
Assessment. This viewpoint is held by some intelligent design proponents, as well as many other people who do not follow ID in particular but who are skeptical of the naturalistic storyline.
My Take. This is an intellectually reasonable position that has much merit. It acknowledges the problems with naturalistic abiogenesis scenarios and the legitimacy of an alternative design explanation. However, it still reposes a (weak) hope for some future discovery that might make the untenable abiogenesis story tenable.
6. “Life almost certainly arose through intelligent guidance or intervention and not solely through natural processes. The very claim of an information-rich, functional system arising through such processes is anathema to our understanding of cause and effect in the world, has never been observed, and is not based on solid scientific footing. Furthermore, there are excellent affirmative reasons to think that life could only have arisen through intelligent guidance or intervention. Although there is no rational reason to think that a plausible naturalistic scenario will ever be discovered, there is value in continuing origin of life research, both because we will learn much about biology in the process and because we will learn more about the hurdles that a naturalistic scenario would have to overcome and the areas in which design is required.”
Assessment. This viewpoint is similar to but stronger than #5, and is perhaps the most common position among intelligent design proponents. It draws a strong, but tentative, inference based on the current state of scientific knowledge, while being open to the general value of additional research.
My Take. In my view, this position is most consonant with the current scientific evidence and, therefore, unsurprisingly, is the view that I personally favor.
7. “Life was created directly by an intelligent being without purely natural processes. To think otherwise is to deny God’s Word, which no amount of scientific undertaking can overturn. Abiogenesis research may yield some peripheral benefits, but as a scientific undertaking on its own merits it is essentially a waste of time and resources.”
Assessment. This viewpoint is occasionally found among some religious adherents. It consists of (i) a declaration of faith, based on a particular interpretation of religious text, coupled with (ii) a certain ignorance of – or perhaps disinterest in – the scientific research.
My Take. Similar to #1, this last position is essentially a faith-based declaration that has little interest in objectively evaluating the evidence. Despite having no wish to denigrate anyone’s religious views, I also have little patience for this position, both because (a) it is unhelpful in assessing the scientific research and moving our knowledge forward, and (b) it is, in my opinion, theologically unsound.
—–
Here is a simple table outlining the above views:
—–
As mentioned, these are broad categories, but are hopefully at least broken down enough to get the thoughts flowing.
Which position most closely resembles your viewpoint? And what additional caveats or clarifications, if any, would you include to more fully represent your position?
—–
UPDATE: Please see comments 87 and 93 for a better approach to viewpoint #7.