John A Davison
A Prescribed Evolutionary Hypothesis
1. Introduction
2. The Prescribed Evolutionary Hypothesis
3. The Indirect Evidence
4. The Direct Evidence
5. Conclusion
Abstract. I propose that phylogeny took place in a manner similar to that of ontogeny by the derepression of preformed genomic information which was expressed through release from latency (derepression) by the restructuring of existing chromosomal information (position effects). Both indirect and direct evidence is presented in support of the Prescribed Evolutionary Hypothesis.
l. INTRODUCTION
Historically there have been two major hypotheses to explain organic change, that of Lamarck, based on the transmission of characters acquired during the life of the individual and that of Darwin, which placed Nature in the role of selecting and thereby preserving those genetic changes which proved to be of advantage to the organism. These changes were presumed to be the means by which evolution proceeded. Each of these hypotheses has been thoroughly tested. The Lamarckian hypothesis was tested by August Weismann in Darwin’s own day with negative results. The Darwinian hypothesis has been tested with limited success. There is no question that artificial selection can significantly alter the phenotype as demonstrated with dogs, goldfish, and a host of other domesticated forms, both plant and animal. Nevertheless, the products of the most intensive selection have not exceeded the species barrier. It seems that sexual reproduction is incapable of transforming species even to new members of the same genus. Even if this could be demonstrated, it seems very unlikely that such a process could ever produce the higher categories of genus, family, order or class. I realize that these are contentious matters and it is with some trepidation that I have abandoned each of these hypotheses in order to offer what seems to me the only real viable alternative. It is the responsibility of the scientist to expose failed hypotheses, but it is equally his responsibility to offer a replacement for them. That is the purpose of this paper. Some of what I will present is not new with me but was proposed long ago by those I will cite, in their own words, so there is no misunderstanding of what they meant.
2. THE PRESCRIBED EVOLUTIONARY HYPOTHESIS
I propose that the information for organic evolution has somehow been predetermined in the evolving genome in a way comparable to the way in which the necessary information to produce a complete organism is contained within a single cell, the fertilized egg. Just as differentiation involves the ordered derepression of pre-existing information, so then I propose, did evolution proceed by a similar means. Viewed in this way, ontogeny and phylogeny become part of the same organic continuum utilizing similar mechanisms for their expression. For those who may be unfamiliar with the history of evolutionary thought, these notions may seem bizarre, but they are in no way original with me. I only propose to extend them somewhat further.
Leo S. Berg in 1922 published his remarkable book, Nomogenesis or Evolution According to Law, in which he presented several examples of what he called phylogenetic acceleration or the premature appearance of advanced features in primitive organisms. Among these were the development of a true placenta in certain sharks (Mustelus laevis), the ciliate protozoon (Diplodinium ecaudatum) in which whole “organ systems†are elaborated within the confines of a single cell, the possession of pneumatic bones in certain flightless reptiles and many other examples of the appearance of advanced features even in organisms for which there is no apparent adaptive significance. Generalizing from several such examples, Berg concluded:
“Evolution is in a great measure an unfolding of pre-existing rudiments.†(Berg [1969] page 406)
In the same volume he quoted William Bateson:
â€ÂFinally, Bateson likewise (1914) inclines to the view that the entire process of evolution may be regarded as ‘an unpacking of an original complex which contained within itself the whole range of diversity which living things present’.†(Berg, page 359).
Pierre Grasse (1977, page 209) reached similar conclusions, apparently independently:
“However that may be, the existence of internal factors affecting evolution has to be accepted by
any objective mind…â€Â
I propose that these internal factors may prove to be the primary if not the sole causes of organic evolution. In short, I suggest that evolution has been largely an emergent process in which the environment may have played, at best, a trivial role. A similar view was expressed by Otto Schindewolf, again independently:
“On the other hand, an unbiased examination of the fossil material itself also reveals that absolutely no direct response to environmental influences or appropriate adaptations in the Lamarckian sense must necessarily be inferred.†(Schindewolf [1993] page 312).
“At most, the environment plays only a similar role with regard to organisms; it can only provoke and set in motion some potential that is already present.” (Schindewolf page 313, his emphasis).
The key words here are already present. Reginald Punnett, in his book Mimicry in Butterflies offered a similar appraisal of the environment (Natural Selection) in 1915:
“Natural selection is a real factor in connection with mimicry, but its function is to conserve and render preponderant an already existent likeness, not to build up that likeness through the accumulation of small variations, as is so generally assumed.†(Quoted in Berg, page 314, my emphasis).
In 1909, Henry Fairfield Osborn presented a similar evaluation of the role for Natural selection:
“In all the research since 1869 on the transformations observed in closely successive phyletic series no evidence whatever, to my knowledge has been brought forward by any paleontologist, either of the vertebrated or invertebrated animals, that the fit originates by selection from the fortuitous.†(Quoted in Berg, page 127).
It should also be noted that Schindewolf was adamant in his refusal to regard evolution as an experimental science.
“Evolution, a unique, historical course of events that took place in the past, is not repeatable experimentally and cannot be investigated in that way.†(Schindewolf, page 311).
As an experimental biologist I was, at first, very reluctant to accept his judgment. In retrospect however, I am now inclined to believe he was correct. If true, it can offer an explanation for the failure of both Darwinism and Lamarckism to provide answers to the causes of evolution. One can hardly expect to demonstrate a mechanism that simply does not and did not exist. We are left with the provisional conclusion that evolution, like ontogeny, has been driven by internal mechanisms the nature of which remain, at present, unknown.
3. THE INDIRECT EVIDENCE
A consideration of evolution as an emergent process suggests a completely new interpretation of certain phenomena. I recounted several examples in an earlier paper (Davison [2000]) so I will only briefly refer to them again. The so-called phenomenon of convergent evolution may not be that at all, but simply the expression of the same preformed “blueprints†by unrelated organisms. Examples include marsupial “molesâ€Â, “wolvesâ€Â, “anteatersâ€Â, “rabbits†(bandicoots), “squirrelsâ€Â, including flying forms (phalangers), “woodchucks†(wombat), â€Âbearsâ€Â, (koala), “mice†(Coenolestes) and most remarkable of all, saber-toothed cats. In Figure 1 Schindewolf presented pictures of the skulls of the marsupial Thylacosmilus atrox opposite that of the placental Eusmilus sicarius, the former from the Pleistocene of Patagonia, the latter from the Oligocene of South Dakota. The two forms are separated by thousands of miles spatially and millions of years temporally. Schindewolf’s legend beneath the figure is of special significance as it bears, not only on the questions raised here, but also, on the whole issue of Intelligent Design with which those questions are clearly related.
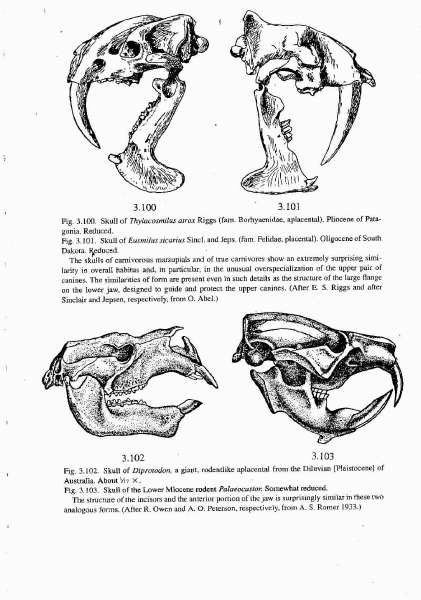
Figure 1
“The skulls of carnivorous marsupials and of true carnivores show an extremely surprising similarity in overall habitus and, in particular, in the unusual overspecialization of the upper pair of canines. The similarities of form are present even in such details as the structure of the large flange on the lower jaw, designed to guide and protect the upper canines.†(Schindewolf, page 261, my emphasis).
Schindewolf also illustrates examples from the plant kingdom of xerophytes from three different plant families all exhibiting the “cactus†phenotype (Cereus Pringlei, Stapelia grandiflora and Euphorbia erosa). In that same plate he compares photographs of the marsupial wolf Thylacinus cynocephalus with the placental wolf Canis lupus. (Schindewolf, plate 5). Similarly, the placental gerbil (Gerbillus) mimics the kangaroo and wallaby both structurally and in mode of locomotion.
It should also be obvious that if specific information was preformed in the evolving genome there would be no need for gradual transformations from one form to another, which remains in accord with the conspicuous absence of transitional intermediates in the fossil record. Furthermore, since such transitional forms are also absent in the contemporary biota, there seems to be no compelling reason to postulate their existence during their evolutionary emergence. In short, evolution may have proceeded by a series of instantaneous transformation (saltations) asindependently proposed by both Schindewolf [1993], Goldschmidt [1940] and more recently by the present author [2004]. Such a mechanism represents the very antithesis of the gradualist Darwinian paradigm. The Darwinian model would explain these phenomena as resulting from the gradual accumulation of mutations. Pierre Grasse commented on the Darwinian view as follows.
“A cluster of facts makes it very plain that Mendelian, allelomorphic mutation plays no part in creative evolution. It is, as it were, a more or less pathological fluctuation in the genetic code. It is an accident on the “magnetic tape†on which the primary information for the species is recorded.†(Grasse, page 243, my emphasis).
One may note that I have used the past tense in reference to evolution. I earlier presented the evidence that macroevolution (true speciation and certainly the formation of the higher categories) is no longer in progress, again a conclusion reached by others long before me, notably Robert Broom. (Davison [2004]).
Note that Grasse uses the present tense in the above quote as well as in the title for his book, The Evolution of Living Organisms. However, in all fairness to Grasse, he also stated:
“The period of great fecundity is over; present evolution appears as a weakened process, declining or near its end. Aren’t we witnessing the remains of an immense phenomenon close to extinction? Aren’t the small variations which are being recorded everywhere the tail end, the last oscillations of the evolutionary movement? Aren’t our plants, our animals lacking some mechanisms which were present in the early flora and fauna?” (Grasse page 71).
Even Julian Huxley, the author of Evolution: the Modern Synthesis, suggested as much in a statement difficult to reconcile with the Darwinian model.
“Evolution is thus seen as a series of blind alleys. Some are extremely short — those leading to new genera and species that either remain stable or become extinct. Others are longer – the lines of adaptive isolation within a group such as a class or subclass. Which run for tens of millions of years before coming up against their terminal blank wall. Others are still longer – the links that in the past led to the development of the major phyla and their highest representatives; their course is to be reckoned not in tens but in hundreds of millions of years. But all in the long run have terminated blindly.†(Huxley [1942] page 571).
This remarkable summary appears just seven pages before the end of the book and would seem to challenge much of what has preceded it.
4. THE DIRECT EVIDENCE
In 1940 Richard B. Goldschmidt [1940] presented the evidence that it is the chromosome, not the gene that is the unit of evolutionary change. While this was not then accepted by the evolutionary establishment, recent karyological studies fully support his perspective. The primary demonstrable differences that distinguish us from our closest primate relatives are revealed in the structure of our chromosomes. They consist of several reorganizations of homologous chromosome segments in the form of translocations, pericentric and paracentric inversions and a single fusion which result in the human complement of 46 chromosomes while the Chimpanzee, Gorilla and Orang each have 48 (Yunis and Prakash [1982]). The important point is that there is no evidence that such transformations involved in any way the introduction of species specific information into the genome. This is further reinforced by the demonstration that we are nearly identical at the DNA level with our close relatives. The simplest explanation is that the information was present in a latent state and simply revealed or derepressed when the chromosome segments were placed in a new configuration (Davison [1993]). In other words, we are dealing with what has been described as “position effects†which also obviously do not involve the introduction of new information from outside the genome. Any change in such gene expressions can only result from the influence of the new structural environment. I find it difficult to imagine how Natural Selection could in any way influence the conditions leading to such chromosome reorganizations. They would seem to have a purely endogenous origin, just as Berg, Bateson and Grasse had suggested long ago. Furthermore, recent studies clearly demonstrate that such chromosome reorganizations do not occur randomly as the Darwinian model would assume. While the technical aspects of their studies are beyond the scope of this paper, the title of the paper, “Hotspots of mammalian chromosomal evolution†indicates, as the authors demonstrate, that there are definitely preferred points at which chromosomes break and recombine. They state in their conclusion section:
“Rather, our analysis supports a nonrandom model of chromosomal evolution that implicates specific regions within the mammalian genome as having been predisposed to both recurrent small-scale duplication and large-scale evolutionary rearrangements.†(Bailey, et al. [2004], my emphasis).
Recent studies (Bauer et al. [2001]) show that genes experimentally introduced near chromosome ends (telomeres) tend to be silenced, this providing further evidence for “position effects†in determining gene expression. While these are developmental phenomena, might they not also have operated during evolution?
Further support for the Prescribed Evolution Hypothesis comes from studies with one of the most primitive of the animal phyla, the Cnidaria (Kortschelt et al. [2003]). Working with the planula stage of the coral Acropora millepora, they found it to be genetically very complex, containing many genes previously thought to be vertebrate innovations. From their summary:
“Acropora millepora provides a unique insight into the unexpectedly deep evolutionary origins of at least some vertebrate gene families.â€Â
The great genetic complexity of a primitive form such as Acropora millepora raises an interesting question concerning the nature of the evolutionary process. Is it not possible that evolution might have involved, to some extent at least, the loss rather than the gain of information? Comparing phylogeny with ontogeny once again, certainly the fertilized egg has a greater developmental potential than the individual cells which result from its division. Thus, just as differentiation results in a progressive loss of potential, so may have phylogeny proceeded in a similar fashion. This perspective also offers a rationale for the irreversibility of the evolutionary process.
5. CONCLUSION
I hope that this brief discussion will serve to stimulate a further investigation into the mystery of organic evolution from a new perspective. The Prescribed Evolutionary Hypothesis (PEH) presents a clear alternative to both the Darwinian and Lamarckian paradigms. It is no longer acceptable to regard evolution as a self-regulating phenomenon guided by random mutation and Natural Selection, especially since it has failed the acid test of experimental verification. PEH is not even a particularly radical idea. Does anyone believe that the laws that are so evident in all of mathematics, physics and chemistry were not somehow prescribed? Science is nothing more than the discovery of that which is there and always has been there. It is interesting to note that Charles Darwin and Alfred Russel Wallace discovered nothing. They had both been profoundly influenced by the writings of Malthus and Lyell and erected their hypothesis largely on those considerations alone, namely extensive periods of time and overpopulation. It is of great significance and much to his credit that Wallace finally abandoned the whole scheme as is so obvious from the title of his last book, The World of Life: A Manifestation of Creative Power, Directive Mind and Ultimate Purpose. (Wallace [1911]). That does not mean that there need to have been supernatural intervention during the process of evolutionary change. Pierre Grasse properly identified the nature of the polarities that continue to inhibit our understanding of the evolutionary process.
“Directed by all-powerful selection, chance becomes a sort of providence which, under the cover of atheism, is not named but which is secretly worshipped. We believe that there is no reason for being forced to choose between “either randomness or the supernatural,†a choice into which the advocates of randomness in biology strive vainly to back their opponents. It is neither randomness nor supernatural power, but laws which govern living things; to determine these laws is the aim and goal of science, which should have the final say. (Grasse, page 107).
Even Albert Einstein, who I don’t believe ever commented on evolutionary matters, offered the following which certainly is not in any sense in conflict with what has been presented here.
“Everything is determined… by forces over which we have no control. It is determined for the
insect as it is for the star.â€Â
Professor Emeritus of Biology, University of Vermont, U.S.A.
REFERENCES
Bailey, J.A., R. Baertsch, W.J. Kent, D. Haussler and E.E. Eichler [2004]. Hotspots of Mammalian Chromosome Evolution. Genome Biology 5: R23.
Bauer, J.A., Y. Zou and W.E. Wright [2001]. Telomere Position Effect in Human Cells. Science 292: 2075-2077.
Berg, L.S. [1969], Nomogenesis or Evolution Determined by Law. M.I.T. Press, Cambridge. (original Russian edition, 1922).
Davison, J.A. [2000], Ontogeny, Phylogeny and the Origin of Biological Information. Rivista di Biologia / Biology Forum 93: 513-523.
Davison, J.A. [2004], Is Evolution Finished? Rivista di Biologia / Biology Forum 97: 111-116.
Goldschmidt, R.B. [1940], The Material Basis of Evolution. Yale University Press.
Grasse, P.P. [1977], Evolution of Living Organisms. Academic Press, New York. (original French edition 1973).
Huxley, J. [1942], Evolution: The Modern Synthesis. Harper, New York and London.
Kortschak, R.D., G. Samuel, R. Saint and D.J. Miller [2003], EST Analysis of the Cnidarian Acropora millepora Reveals Extensive Gene Loss and Rapid Sequence Divergence in the Model Invertebrates. Current Biology 13: 2190-2195.
Schindewolf, O. [1993], Basic Questions in Paleontology. The University of Chicago Press, Chicago and London. (original German edition, 1950).
Wallace, A.R. [1911], The World of Life: A Manifestation of Creative Power, Directive Mind and Ultimate Purpose. Moffat Yard and Co., New York.
Yunis, J.J. and O. Prakash [1982], The Origin of Man: A Chromosomal Pictorial Legacy. Science 215: 1525-1530.