Imagine this paradoxical scenario: NASA decides to construct a new space station and a new space shuttle. It assigns to a team of engineers the job of designing the station and to another team the job of designing the shuttle, without any specifications about the relations between the two projects and the mandate that they must not exchange information (the projects are top secret). The teams finish their work and the systems are launched into the space. Would you bet one cent that the shuttle will succeed to dock to the station to exchange materials and astronauts? Sure not because when two complex systems must interface in a complex manner (as the station and the shuttle when they are coupled together) their projects have to be thought together just from the beginning. Eventually two different teams can be in charge of the two designs but the teams must strictly collaborate and exchange a lot of information.
This Kafkian story introduces us to the concept of what could be called “coupled complex specified information” (cCSI), that is a particular form of complex specified information (CSI) concerning two systems instead of one alone. In general when two systems must share a complex interface the CSI of the interface is strictly correlated to the CSI of the systems. As such the three projects cannot arise independently but have to be developed almost together with a higher direction that considers them almost like a unique system. For a symbolic representation of the cCSI between two systems A and B see the following figure:
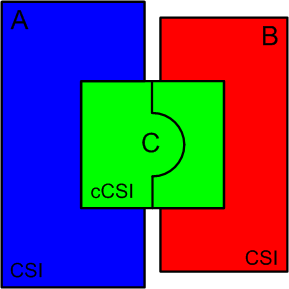
Of course, beyond the above astronautic example, one can find in technology many other more or less simple examples of cCSI. In mechanics: a bolt and a nut; a lock and its key. In railway: the train and the railroad; the locomotive and the wagon. In informatics: the network interface between two computers. In electronics: a connector and its plug or in general the interface between two devices. In communication: transmitter and receiver must share communication protocols. In language: the horizontal and vertical words in a crossword puzzle. I leave the reader the job of finding other examples of complex matching between systems.
The examples of communication and networking leads us to emphasize that an interface (as in general any complex system) may be composed of a material part (as the adapter card) and an abstract part (as the protocols). When information processing is involved these two aspects (hardware and software) are always present. Therefore in this case also the cCSI of the interface has to be considered from these two points of view.
The cCSI systems necessarily are intelligently designed. In fact to develop them a supervisor is necessary who knows in the same time the two systems and their interface. Only intelligence can be such supervisor because only intelligence is able to achieve knowledge sharing. The first premise of knowledge sharing is memory. Naturalistic and mechanic processes as the evolutionary ones just fail this premise since have no memory and thus are unable to manage correlated parallel jobs, where one must know and recall how A and B are made to create C that must interface both.
What about biology and cCSI? In molecular biology examples of matching systems are enzymes, which are catalytic proteins having a unique very specific 3-D shape that must fit their target substrate like a key fits in a lock. In morphological biology indeed there are spectacular examples of cCSI. One for all, consider the sexual reproductive organs in mammals. They must match at the every level of the biological stack: from bio-chemistry to tissues, from genetics to morphology. Last but not least also the behavioral level is involved if two individuals of different sex have to arrive to mating and generating off-spring. Hence also the behaviors belong to the cCSI of interfacing. About the matching between behavioral patterns and physical forms as sign of intelligent design see here .
It is unimaginable that reproduction and genitals arose by Darwinian evolution (that is for random mutations and natural selection). First, as a matter of principle: evolution needs reproduction; without reproduction no evolution. Therefore how can reproduction be the effect of evolution if evolution is an effect of reproduction? It’s an impossible causality inversion. Second, for a technical reason: how could the male organs arise independently from the female organs given the cCSI they share? In fact the Darwinian processes work in the single individual. They are blind and unaware of the processes running in other individuals. Random mutations that happen in a genome have nothing to do with the mutations in another one.
To sum up, biological cCSI cannot arise by evolution because the Darwinian independent processes would be as the above two NASA engineering teams that don’t speak together. In both cases the products, independently made, will never match. Darwinian evolution of the cCSI apparatuses is even more unbelievable than the above NASA story.
About cCSI and its refutation of a Darwinian explanation of sex and reproduction see also here .